发布时间:2020年03月27日 10:01:06 来源:振东健康网
张静,李素芳,陈红△,宋俊贤
(北京大学人民医院心血管内科,急性心肌梗死早期预警和干预北京市重点实验
室,北京大学人民医院心血管转化医学研究中心,北京100044)
【摘要】目的:评估miR-106b-5p对内皮细胞基因表达谱的影响。方法:对动脉粥样硬化组(n=9)及对照组(n=9)血浆中miRNAs表达谱进行筛查,采用共表达网络分别对两组样本全体miRNAs的共表达模式进行分析。选取从两组网络中得到的共表达地位差异最为显著的miR-106b-5p进一步研究,通过转染miR-106b-5pmimics上调人脐静脉内皮细胞的miR-106b-5p表达水平,筛查转染后差异基因表达谱,并进一步通过日本京都基因与基因组百科全书(KyotoEncyclopediaofGenesandGenomes,KEGG)信号转导通路数据库对差异基因富集的信号通路进行分析。结果:动脉粥样硬化组患者血浆中miRNAs的共表达模式(140个节点,1154条连接线)与对照组(140个节点,612条连接线)相比存在明显差异,在过表达miR-106b-5p后人脐静脉内皮细胞有746个基因水平发生了显著变化(组间差异倍数≥1.5,芯片错误发现率<0.01),主要包括磷脂酰肌醇-3激酶(phosphoinositide3-kinase,PI3K)/蛋白激酶B(proteinkinaseB,PKB,又称Akt)信号通路、哺乳动物雷帕霉素受体蛋白(mammaliantargetofrapamycin,mTOR)信号通路、转化生长因子-β(transforminggrowthfactor-β,TGF-β)信号通路、酪氨酸激酶-信号转导及转录激活因子信号通路(januskinase/signaltransducerandactivatoroftranscription,Jak-STAT)信号通路、肿瘤坏死因子(tumornecrosisfactor,TNF)信号通路、toll样受体(toll-likereceptor,TLR)信号通路、血管内皮生长因子(vascularendothelialgrowthfactor,VEGF)信号通路等20个信号通路。结论:动脉粥样硬化患者血浆中miRNAs共表达模式发生了显著变化,其中共表达地位差异最为显著的miR-106b-5p可靶向调节血管内皮细胞多个信号通路。
【关键词】动脉粥样硬化;微RNAs;转录组;内皮细胞
[中图分类号]R541.4
Role of miR-106b-5p in the regulation of gene profiles in endothelial cells
ZHANG Jing,LI Su-fang,CHEN Hong△ ,SONG Jun-xian
(Department of Cardiology, Peking University People’s Hospital; Beijing Key Laboratory of Early Prediction and Intervention of Acute Myocardial Infarction,Peking University People’s Hospital; Center for Cardiovascular Translational Research, Peking University People’ s Hospital, Beijing 100044, China)
【ABSTRACT Objective】: To evaluate the role of miR-106b-5p in the regulation of gene expression in endothelial cells. Methods: The Taqman low-density microRNAs (miRNAs) array (TLDA) was used to identify miRNA expression profiles in the plasma of patients with atherosclerotic coronary artery disease (CAD) (atherosclerosis group, n=9) and individuals without atherosclerotic CAD disease (control group, n=9).A weighed and undirected miRNA coexpression network analysis was performed to investigate the interactions among miRNAs in the two groups. MiR-106b-5p, whose coexpression pattern in atherosclerosis group was most different from that of control group, was further studied. Human umbilical vein endothelial cells (HUVEC) were transfected with miR-106b-5p mimic or negative control mimic, and Affymetrix GeneChip Human Transcriptome Array 2.0 was used to screen the differential gene expression profiles after transfection. And the signal transduction pathway of differential gene profiles was further analyzed in Kyoto Encyclopedia of Genes and Genomes (KEGG) signal pathway database. After parsing the whole KEGG database,all differentially expressed genes involved pathways were extracted, and the hypergeometric distribution was used to calculate the pathway enrichment. Results:The coexpression pattern of the patients with atherosclerosis (140 nodes, 1 154 edges)
differed from that of the non-atherosclerosis control group (140 nodes, 612 edges).The analysis of array data with Significant Analysis of Microarray (SAM) identified 746 significantly deregulated genes (fold change ≥ 1.5 and false discovery rate <0.01) altered by overexpression of miR-106b-5p with miR-106b-5p mimic in HUVEC.By calculating the pathway enrichment, we found that multiple signaling pathwaysenriched in differential gene profiles were closely related to the process of formation and rupture of atherosclerotic plaque, including phosphatidylinositol-3 kinase (PI3K)/3protein kinase B (PKB, also called Akt), mammalian target of rapamycin (mTOR),transforming growth factor-β (TGF-β), janus kinase / signal transducer and activator of transcription (Jak-STAT), tumor necrosis factor (TNF), toll like receptor (TLR) and hypoxia-inducible factor 1α (HIF-1α) and other signal pathways. Conclusion: The coexpression pattern of miRNAs in plasma of patients with atherosclerosis is more significantly changed than that of individuals without atherosclerotic disease.MiR-106b-5p, which shows the most significant difference between groups, targetsmultiple signal pathways in vascular endothelial cells, and might play an important role in the regulatory network of atherosclerotic gene expression.
【KEY WORDS】 Atherosclerosis; MicroRNAs; Transcriptome; Endothelial cells
microRNAs(miRNAs)是一类在进化上高度保守的非编码小分子单链 RNA(约 22 nt),在转录后水平负性调控基因表达 [1] 。目前在人体中已发现 1 000 多个 miRNAs,约 30%的人类蛋白编码基因受到 miRNAs 的调控 [2] 。我们以往的研究表明,冠状动脉(冠脉)粥样硬化性心脏病患者血浆中存在包括 miR-106b-5p等在内的多种 miRNAs 水平显著升高 [3] ,也有研究发现 miR-106b-5p 在脑卒中、癌症等疾病进程中发挥了重要的调控作用,通过靶向调节特定的靶基因可以调控细胞炎症反应、氧化应激、凋亡等重要环节 [4-5] 。
共表达网络分析是一种基于 miRNAs 整体网络背景下,系统分析每对miRNAs之间相互关系以及具体某个 miRNAs在整体网络中的重要程度和贡献性的生物信息学技术 [6] 。
本研究拟将动脉粥样硬化疾病患者与对照者血浆中 miRNAs 表达谱进行比对,并进一步选取从两组样本的共表达网络得到的共表达地位差异最为显著的miRNA,进一步探讨目标 miRNA 对血管内皮细胞基因表达谱的调节作用。
1 资料与方法
1.1 研究对象的入选和排除标准
选择2011年12月至2013年10月于北京大学人民医院心脏中心心内科住院并行冠脉造影检查的冠脉粥样硬化性心脏病患者,包括不稳定性心绞痛(unstable angina,UA)以及非心源性胸痛对照者。UA 患者的诊断标准参照 2011 年美国心脏病学会(American College of Cardiology,ACC)/美国心脏协会(American Heart Association,AHA)关于 UA/非 ST 段升高心肌梗死(non-ST-elevation myocardial infarction,NSTEMI)的诊断和治疗指南。所有非心源性胸痛患者(对照组)经冠脉造影检查均未发现冠脉管腔狭窄,同时在入院之前未曾服用过他汀类药物,并经过随访明确其胸痛的原因为非心源性,如胃食道返流、心脏神经症等其他原因。
排除标准包括:各种急慢性炎症、肝肾功能异常、自身免疫性疾病、恶性肿瘤以及血液系统疾病等。获取入选者主要临床资料,包括:年龄、性别、身高、体重、血压、腰围、臀围、临床血生化指标、高血压病史、糖尿病病史、高脂血症病史、吸烟状况及服用药物的情况等。
1.2 血浆中 miRNAs 的 提取和芯片检测
所有研究对象的血浆均在患者接受冠脉造影术之前抽取,使用EDTA抗凝管保存于4 ℃冰箱,于1 h内在4 ℃下分离血浆。参照文献[3]中的方法,采用miRNAs提取试剂盒(miRNeasy kit,Qiagen,德国)提取血浆中含有miRNAs的总RNAs,并进行miRNA反转录实验(A、B板分别采用15 ng RNA起始)。采用美国ABI公司的反转录试剂盒(TaqMan® MicroRNA Reverse Transcription Kit)和miRNAs特异的反转录引物池(Megaplex™ RT Primer Pools, Human Pools Set v3.0)进行miRNAs的反转录反应 [3] 。随后采用预扩增试剂(TaqMan ® PreAmp Master Mix,2×)和miRNAs特异的Megaplex预扩增引物池(Megaplex™ PreAmp Primer Pools,Human Pools Set v3.0)进行miRNAs的预扩增反应 [3] ,采用美国ABI公司的Taqman 低密度芯片(Taqman low-density array,TLDA)(TaqMan Array Human MicroRNA A+B Cards Set v3.0)进行筛查实验,采用Mamm U6作为内参,目标miRNAs的相对表达水平表示为2 -(CT[miRNA]-CT[U6]) 。两组间差异倍数的计算采用2 -∆∆Ct 的方法 [3] 。
1.3 共表达网络分析
采用 miRNAs 共表达网络分析(coexpression of miRNAs network analysis)进一步在整体 miRNAs 网络的背景下对 miRNAs 的表达模式进行分析,使用Cytoscape v.3.5.0 软件分别对动脉粥样硬化患者组、对照组的 miRNAs 对进行共表达分析,得到疾病组和对照组各自的静态共表达整体网络 [7] 。
1.4 人脐静脉内皮细胞培养及 miRNA mimic 转染
人脐静脉内皮细胞(human umbilical vein endothelial cells,HUVEC)获取于北京大学基础医学院。细胞接种于 6 孔板,待细胞融汇率达 70%~80%,换新的全培养液(含 P/S 双抗和血清)2.5 mL,进行 miRNA mimic(或隐性对照 negative control mimic)的转染,终浓度为 30 nmol/L,转染所用试剂为 Lipofectamine RNAiMAX Transfection Reagent(Invitrogen,美国),详细方法参照文献[8]。转染 24 h 后,用磷酸盐缓冲液清洗 6 孔板 3 遍,采用 TRIzol 法提取细胞总 RNA。以实时聚合酶链反应(real-time polymerase chain reaction,real-time PCR)实验检测转染后目标 miRNA 水平升高的倍数,采用 miRNAs PCR 预混试剂(TaqMan Universal PCR Master Mix,No AmpErase® UNG)及预设计的 FAM 染料标记miRNA MGB 荧光探针(Taqman Small RNA Assay,20×)。
1.5 基因表达谱芯片筛查及 信号通路富集性分析
对进一步采用 Affymetrix GeneChip Human Transcriptome Array 2.0 芯片对转染后的基因表达谱进行检测,使用微阵列显著性分析(significant analysis of microarray,SAM)软件包筛选差异表达的基因,通过设置错误发现率(false discovery rate,FDR)<0.01,差异倍数(fold change)≥1.5,得到组间差异表达的基因。进一步通过日本京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)的信号通路数据库对差异基因富集的信号通路进行分析 [9-10] ,构建基因间的相互作用网络 [9-11] 。
1.6 统计学分析
数值型变量以均数±标准差表示,两组间差异性比较采用 Student’s t 检验;分类型变量采用百分率(%)表示,两组间差异性比较采用卡方(χ 2 )检验或 Fisher精确概率法(Fisher’s exact test)。对双变量呈正态分布的连续性变量,采用 Pearson线性相关分析,对双变量呈非正态分布的等级变量,采用 Spearman 等级相关分析。由 SPSS13.0 统计软件完成,以 P<0.05 为差异有统计学意义。
2 结果
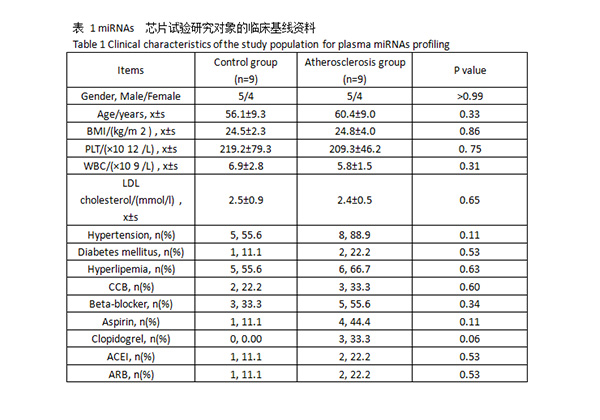
2.1 MiRNAs 芯片试验 研究对象的临床基线资料
共入选研究对象 18 例,其中包括冠脉粥样硬化性心脏病患者(动脉粥样硬化组,n=9)及经冠脉造影检查不存在冠脉粥样硬化症的非心源性胸痛患者(对照组,n=9)。对于芯片筛查对象,两组间各项临床基线资料(包括年龄、性别、体重指数、临床血生化指标、高血压病史、糖尿病病史及高脂血症病史等)差异无统计学意义(P>0.05,表 1)。
2.2 miRNAs 芯片 共表达网络分析
对动脉粥样硬化组及对照组的血浆 miRNAs 进行 Taqman miRNAs TLDA 低密度芯片(A+B 板)检测,并根据两组标本中 miRNAs 的表达水平的相关性分别建立共表达网络。在共表达网络中,每一个节点(node)代表一个 miRNA,而节点与节点之间的连接线(edge/link)代表两个 miRNAs 之间存在表达水平线性相关关系。通过对共线性表达关系较强的 miRNAs 对(皮尔逊相关系数≥0.75,P<0.05,FDR<0.01)构建共表达网络,发现动脉粥样硬化组患者血浆中 miRNAs的共表达模式(140 个节点,1 154 条连接线)与对照组(140 个节点, 612 条连接线)相比存在明显差异(图 1A、B)。
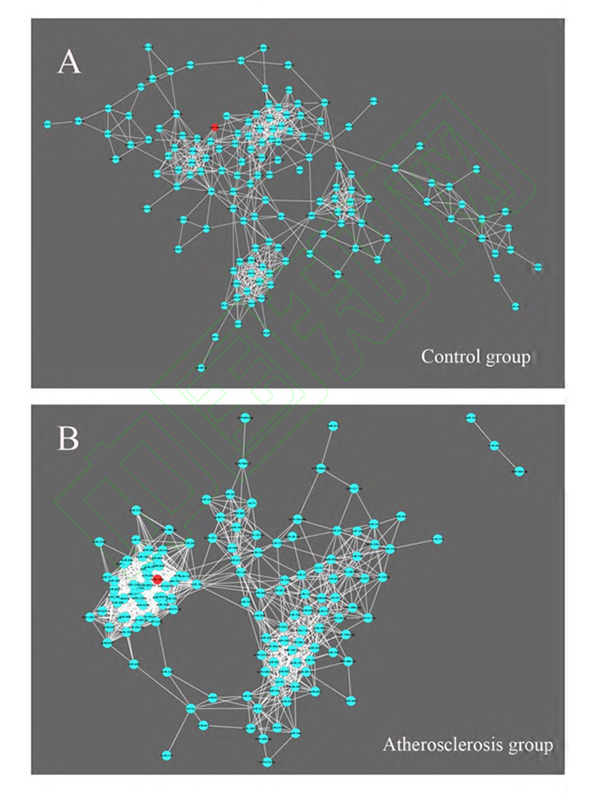
图 1 对照 组(A ) 与 动脉粥样硬化 组(B ) 血浆中 miRNAs 的共表达网络分析图
Figure 1 Coexpression network analysis of plasma miRNAs in controls (A) and atherosclerosis patients (B)
因两个网络中所含的 miRNA 数量不同且表达关系也不同,为了进行更严谨的横向比较,本研究进一步对同一 miRNA 在不同网络中的 miRNA 值进行了标准化后再比较,标准化方法为将某一 miRNA 的连接度(degree)值除以该网络中连接度最大值[Normalized degree(i)=Degree(i)/Degree(Max)]。然后比较同一miRNA 在不同网络中标准化后连接度的差值(DiffK)以及该差值的绝对值(|DiffK|),来体现目标 miRNA 在不同网络中变化的程度(表 2) [7] 。本研究发现,miR-106b-5p 在两组间共表达网络地位差异最为显著,|DiffK|值为 0.78。

2.3 过表达 miR-106b-5p 对 对 HUVEC 基因表达谱 的 影响
选取从两组样本的共表达网络得到的共表达地位差异最为显著的miR-106b-5p 进行进一步研究。转染 miR-106b-5p mimics 后 24 h,人脐静脉内皮细胞的 miR-106b-5p 表达水平发生显著上调(组间差异倍数=1 401.2,P<0.001)。
采用高通量人基因表达谱芯片 Affymetrix GeneChip Human Transcriptome Array 2.0 筛查转染过表达 miR-106b-5p 后 HUVEC 细胞发生显著变化的 mRNA表达谱,发现在过表达 miR-106b-5p 后,聚类分析的热图显示 HUVEC 细胞有 746个基因水平发生了显著变化(组间差异倍数≥1.5,芯片 FDR<0.01,图 2),绝大多数为下调表达的负性调节作用(253 个基因上调表达,493 个基因下调表达)。
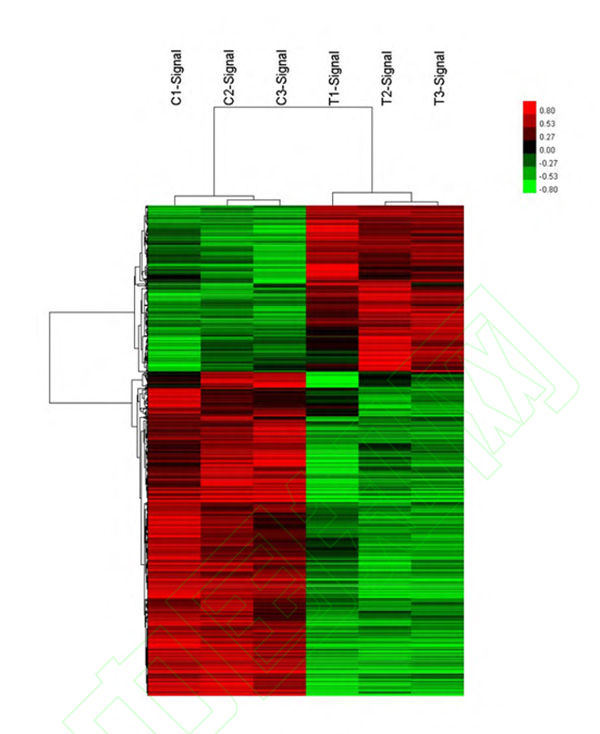
图 2 转染 miR-106b-5p mimics 后人脐静脉内皮细胞基因表达谱发生显 著变化Figure 2 Profile of mRNAs in human umbilical vein endothelial cells transfected with miR-106b-5p mimics compared with negative control mimic
2.4 差异表达基因的 信号通路富集性分析
进一步通过 KEGG 信号转导通路数据库对组间差异表达基因富集的信号通路进行信号通路富集性分析(pathway enrichment analysis)。目前,KEGG 是有关信号通路的主要公共数据库 [9] 。根据挑选出的差异基因,计算这些差异基因同信号通路的超几何分布关系,信号通路富集性分析会对每个有差异基因存在的信号通路返回一个 P 值,P 值具有显著性(P<0.05)表示组间差异表达基因在该信号通路中出现了富集 [10] 。同时基于 KEGG 信号转导通路数据库,还可寻找不同样品组间的差异表达基因可能参与调控的细胞通路,同时将上下调的差异基因映射到每一个信号通路地图(pathway map)上,定位关键通路的关键基因 [10-11] 。
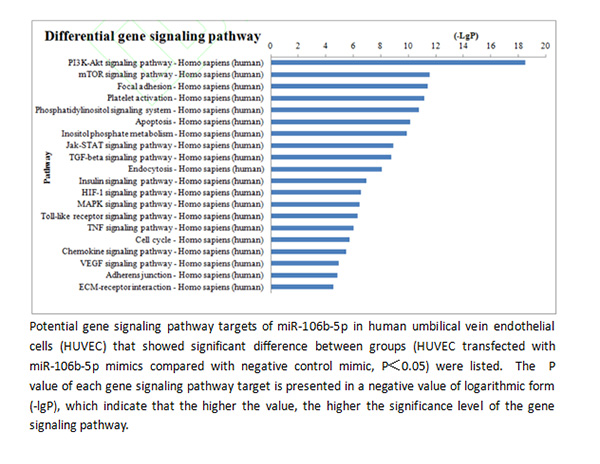
通过信号通路富集性分析发现,在过表达 miR-106b 前后,人脐静脉内皮细胞 HUVEC 表达水平发生显著性变化的这部分基因主要在 20 个信号通路中出现了显著性富集(P<0.05,图 3),其中有多个与动脉粥样硬化斑块形成过程、由稳定状态向不稳定状态转化的过程密切相关的信号通路,包括磷脂酰肌醇-3 激酶(phosphoinositide 3-kinase,PI3K)/蛋白激酶 B(protein kinase B,PKB,又称Akt)信号通路、哺乳动物雷帕霉素靶向基因(mammalian target of rapamycin,mTOR)信号通路、转化生长因子-β(transforming growth factor-β,TGF-β)信号通路、酪氨酸激酶-信号转导及转录激活因子信号通路(janus kinase/ signal transducer and activator of transcription,Jak-STAT)信号通路、肿瘤坏死因子(tumor necrosis factor,TNF)信号通路、toll 样受体(toll-like receptor,TLR)信号通路、血管内皮生长因子(vascular endothelial growth factor,VEGF)信号通路及缺氧诱导因子(hypoxia-inducible factor 1α,HIF-1α)等信号通路(图 3)。
本研究进一步基于 KEGG 数据库,利用 KEGG 数据库中的基因之间、基因产物之间的作用关系,通过数据库搜索得到每个基因与其他基因的作用关系,发现目标基因群之间作用关系,突破单一的信号通路,在整个数据库中定位上游蛋白和下游蛋白,而后构建基因间的全局信号转导网络(global signal transduction network)(图 4) [10-11] 。从该网络图中可以看出,miR-106b-5p 的多个靶基因,包括 PI3K、AKT/PKB、JAK、TGF-β 和磷酸酶-张力蛋白同源物(phosphatase and tensin homolog,PTEN)等连接度>10 的基因在该信号转导网络中均起到了非常重要的作用。
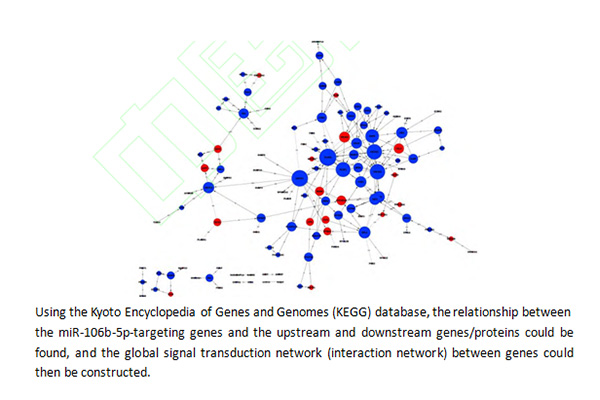
图 4 人脐静脉内皮细胞中 中 miR-106b-5p 靶向调控基因及其上、下游基因所构建的全局信号转导网络
Figure 4 Global signal transduction network constructed with potential gene targets of miR-106b-5p and the upstream and downstream genes in human umbilical vein endothelial cells
3 讨论
一系列研究提示血管内皮细胞损伤是动脉粥样硬化疾病发生的始动因素 [12] ,而 miRNAs 通过参与调控血管内皮细胞的功能,从而影响动脉粥样硬化的发生和发展,加速斑块由稳定状态向不稳定状态转化 [13] 。我们以往的研究表明,不稳定性冠状动脉粥样硬化性心脏病患者血浆中多种 miRNAs 水平显著升高,且其升高的原因可能是由于疾病状态下血管内皮细胞源性的微颗粒数量的显著增加导致的 [3] ,这些在患者血浆中水平升高的 miRNAs 可能来自于血管内皮细胞的释放,且在动脉粥样硬化疾病状态下血管内皮细胞中 miR-106b-5p 水平反而会发生降低 [3] 。
共表达网络分析是一种系统分析每对 miRNAs 之间相互关系的方法,这种将单个miRNAs的水平改变置于整体miRNAs共表达相关网络背景之下的分析方法优于以往专注于单个 miRNA 水平上调还是下调的传统分析方法 [6, 14] 。本研究进一步采用了共表达网络分析对全体 miRNAs 的共表达模式进行了系统分析,发现动脉粥样硬化患者血浆中的 miRNAs 不仅表达水平发生了显著变化,而且动脉粥样硬化组共表达网络所包含的连接线数量(miRNAs 之间的共表达关系)要远远高于对照组,这种有趣的现象可能再次揭示了 miRNAs 在疾病状态下所发挥的协同调控作用。
之后,本研究根据 miRNAs 在动脉粥样硬化组样本与对照组样本的两个网络中共表达地位的差异情况,发现 miR-106b-5p 的|DiffK|值为整个共表达网络中最高值,在两组间共表达网络地位差异最为显著,从生物信息学角度分析,相较于网络中其余的 miRNAs,miR-106b-5p 所处的位置对于全体共表达网络具有更重要的调控价值,对于维系网络的稳定性处于极端重要的地位 [6, 14] 。以往研究发现,miR-106b-5p 可通过靶向调节髓细胞白血病-1(myeloid cell leukemia-1,MCL-1)基因的表达水平,从而在缺血性脑卒疾病进程中抑制细胞凋亡和氧化应激反应,可能是缺血性脑卒中疾病的潜在治疗靶点 [4] 。此外,miR-106b-5p 还可调节 PI3K/ AKT 信号通路,从而在乳腺癌疾病进程中发挥重要调控作用 [5 ] 。 因此,本研究选择 miR-106b-5p 作为进一步离体血管内皮细胞试验的研究对象,发现转染 miR-106b-5p mimics 后,血管内皮细胞的基因表达谱发生了显著变化。
本研究进一步通过 KEGG 信号转导通路数据库对组间差异表达基因所靶向调控的信号通路进行信号通路富集性分析,发现在过表达 miR-106b-5p 前后,HUVEC 细胞表达水平发生显著性变化的这部分基因主要在 20 个信号通路中出现了显著性富集。在出现显著性富集的这部分信号通路中,有多个与动脉粥样硬化斑块形成过程、由稳定状态向不稳定状态转化的过程密切相关的信号通路,涉及炎症、凋亡、细胞趋化、血管新生以及 DNA 损伤修复等多方面作用 [15-18] ,提示 miR-106b-5p 可通过调控血管内皮细胞功能的多个关键因素影响动脉粥样硬化斑块的形成及破裂过程。
本研究发现 miR-106b-5p 是 PI3K/AKT/PTEN 信号通路的重要调控因子,与以往研究一致 [5, 8] ,可能通过抑制内皮细胞凋亡、抑制炎症因子释放等方式在动脉粥样硬化疾病起始阶段发挥保护血管内皮细胞作用 [15-16] 。此外,本研究发现miR-106b-5p 是 TGF-β 信号通路的关键调节因素,可靶向 TGF-β 信号通路中多个关键性蛋白,而 TGF-β 信号通路正是决定炎症和纤维化平衡从而影响斑块不稳定性的关键性信号通路 [17-18] 。miR-106b-5p也可靶向调节 Jak-STAT 信号通路、TNF信号通路和 toll 样受体信号通路等重要炎症反应信号通路,可能从多个方面拮抗血管内皮细胞的炎症反应。此外,miR-106b-5p 还可能通过调节血管新生相关的信号通路(VEGF 信号通路、HIF-1α 等信号通路)影响动脉粥样硬化斑块的稳定性。
本研究存在以下不足:(1)本研究仅选择了在两组间共表达网络地位差异最为显著的 miR-106b-5p 进行研究,其他在组间共表达地位存在显著差异的miRNAs 的作用有待进一步研究。(2)本研究采用了表达谱芯片筛查和生物信息学预测的方法对 miR-106b-5p 的功能进行了初步研究,有必要通过体内和体外实验证实 miR-106b-5p 在动脉粥样硬化斑块形成和破裂过程中的作用和分子机制。
总之,本研究发现动脉粥样硬化患者血浆中 miRNAs 共表达模式发生了显著变化,其中共表达地位差异最为显著的 miR-106b-5p 可靶向调节血管内皮细胞多个基因表达,为基于 miR-106b-5p 的治疗策略向临床应用转化提供了参考,其功能及潜在的分子治疗价值值得进一步研究。
参考文献
[1]Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function [J].Cell, 2004, 116(2): 281-297.
[2]Small EM, Olson EN. Pervasive roles of microRNAs in cardiovascular biology [J].Nature, 2011, 469(7330): 336-342.
[3]Ren J, Zhang J, Xu N, et al. Signature of circulating microRNAs as potential biomarkers in vulnerable coronary artery disease [J]. PLoS One, 2013, 8(12): e80738.
[4]Li P, Shen M, Gao F, et al. An antagomir to microRNA-106b-5p ameliorates cerebral ischemia and reperfusion injury in rats via inhibiting apoptosis and oxidative stress [J]. Mol Neurobiol, 2017, 54(4): 2901-2921.
[5]Li N, Miao Y, Shan Y, et al. MiR-106b and miR-93 regulate cell progression by suppression of PTEN via PI3K/Akt pathway in breast cancer [J]. Cell Death Dis, 2017,8(5): e2796.
[6]Wang J, Haubrock M, Cao KM, et al. Regulatory coordination of clustered microRNAs based on microRNA-transcription factor regulatory network [J]. BMC Syst Biol, 2011, 5(12): 199.
[7]Prieto C, Risueño A, Fontanillo C, et al. Human gene coexpression landscape:confident network derived from tissue transcriptomic profiles [J]. PLoS One. 2008,3(12):e3911.
[8]Zhang J, Li SF, Chen H, et al. MiR-106b-5p inhibits tumor necrosis factor-α-induced apoptosis by targeting phosphatase and tensin homolog deleted on chromosome 10 in vascular endothelial cells [J]. Chin Med J (Engl), 2016,129(12):1406-1412.
[9]Ogata H, Goto S, Sato K, et al. KEGG: Kyoto Encyclopedia of Genes and Genomes [J]. Nucleic Acids Res, 1999, 27(1): 29-34.
[10]Du J, Yuan Z, Ma Z, et al. KEGG-PATH: Kyoto encyclopedia of genes and genomes-based pathway analysis using a path analysis model [J]. Mol Biosyst, 2014,10(9): 2441-2447.
[11]Kanehisa M, Goto S, Sato Y, et al. KEGG for integration and interpretation of large-scale molecular data sets [J]. Nucleic Acids Res, 2012, 40 (Database issue):D109-114.
[12]Ross R. Atherosclerosis: an inflammatory disease [J]. N Engl J Med, 1999,340(2): 115-126.
[13]任景怡, 许宁, 韩冠平, 等. microRNAs 参与动脉粥样硬化疾病的发生[J].中国生物化学与分子生物学报, 2011, 27(6): 511-515.
[14]Zampetaki A, Willeit P, Drozdov I, et al. Profiling of circulating microRNAs:from single biomarkers to re-wired networks [J]. Cardiovasc Res, 2012, 93(4):555-562.
[15]Weber C, Noels H. Atherosclerosis: current pathogenesis and therapeutic options [J]. Nat Med, 2011, 17(11): 1410-1422.
[16]Zernecke A, Weber C. Chemokines in the vascular inflammatory response of atherosclerosis [J]. Cardiovasc Res, 2010, 86(2): 192-201.
[17]Lutgens E, Daemen MJ. Transforming growth factor-β: a local or systemic mediator of plaque stability? [J]. Circ Res, 2001, 89(10): 853-835.
[18]Lutgens E, Gijbels M, Smook M, et al. Transforming growth factor-beta mediates balance between inflammation and fibrosis during plaque progression [J]. Arterioscler Thromb Vasc Biol, 2002, 22(6): 975-982.


移动平台
微信平台
